4 Mutation And Genetic Load
Ms. Shalini Singh and Prof. GK Kshatriya

TABLE OF CONTENTS
1. Introduction
2. General characters
3. Stages
4. Types
i. Morphological Mutations
ii. Lethal Mutation
iii. Biochemical Mutation
iv. Resistant Mutation
v. Spontaneous Mutation
vi. Conditional Mutation
vii. Point Mutation
viii. Induced Mutation
5. Mutation :Biochemical Level
6. Mutation : Molecular Level
7. Effect on Nucleotide sequences
i. Chemical mutations
ii. Alkylating Agents
iii. Acridine Dyes
8. Deleterious Mutations
i. Dominant Mutation as a cause of disease
ii. Chromosomal aberration
9. Genetic load: Definition and Concepts
10. Causes
i. Mutation
ii. Segregation
iii. Inbreeding
iv. Sexual Reproduction
v. Migration
11. Case Studies
1. Introduction
Molecular Anthropology is a field of anthropology which is used to determine evolutionary links between ancient and modern human population as well contemporary species by examining the DNA sequence in different population in order to establish the closeness of relationship between populations. The variations are also pivotal in tracing the migration and settlement pattern. Human genome is not static and is subjected to variety of heritable change as without the occurrence of hereditary changes life forms could have not been evolved and diversified. Alternate form of gene can be resulted due to sudden heritable change in the sequence. Mutation can be defined as the abrupt change in the genotype of an organism that is not a result of recombination and includes all types of heritable genetic change of an organism. The process by which mutation is introduced in a gene is termed as mutagenesis whereas a resulted phenotype with mutation is called a mutant. Mutation is the ultimate source of all genetic variation and provides the raw material for evolution. It results from formation of allele where new organisms will be able to evolve and adapt to environmental change. Thus mutation is used to designate processes by which hereditary changes arise
General characters of mutation
- Mutations are generally recessive but dominant mutation also occurs.
- It is harmful to an organism.
- Mutation is random and occurs at any time and in any cell of an organism.
- Mutation being recurrent as the same mutation can occur again and again.
Stages at which mutations occur:
Mutation can occur at any stage of development only if the situation is met:
Mutation in germinal cell: Before differentiation, mutation in the germinal cell occurs in germinal and also influences several gametes and further influencing all the individuals derived from them.
Mutation in gamete or zygote: In this stage a single individual will carry the mutation.
Mutation in a cell: In this stage the mutation occurs after the zygote has undergone one or more division, here only a part of body will show the mutant character. The mutation at this stage is called somatic mutation.
Types of Mutation: Mutation can be classified on the method of its detection, the main types are
I. Morphological Mutations
II. Lethal Mutation
III. Biochemical Mutation
IV. Resistant Mutation
V. Spontaneous Mutation
VI. Conditional Mutation
VII. Point Mutation
VIII. Induced Mutation
Morphological Mutation
It affects the outward appearance of an individual including colour, shape size etc.
Lethal Mutation
It involves genotypic changes leading to death of an individual and can be easily studied for mutation frequencies
Biochemical Mutation
These types of mutations are identified by a deficiency so that a defect can be rectified by supply of that deficient nutrient in the mutant.
Resistant Mutation
This type of mutation is identified by their ability to grow in the presence of a pathogen or antibiotic.
Conditional Mutation
This type of mutation allows the mutant phenotype to express under permissive conditions.
Point Mutation:
A point mutation causes replacement of single base nucleotide with another base nucleotide in a genetic material. Point mutation can be of three types:
- Missense Mutation- A type of mutation that alters the codon and specifies a different amino acid is known as Missense Mutation
- Nonsense Mutation- A type of mutation where sequence of DNA results in premature stop codon.
- Stop Mutation- A type of mutation which results in no change of amino acid or its functionality when the altered or changes messenger RNA is translated.

Figurehttp://upload.wikimedia.org/wikipedia/commons/6/69/Point_mutations-en.png Spontaneous Mutation
The type of mutation that occurs without treatment of organism with an exogenous mutagen and it is the ultimate source of natural genetic variation in populations. Spontaneous mutation arises from a variety of sources including errors in DNA replication, transposition of transposable genetic elements and spontaneous lesions.
Induced Mutation
These type of mutation arise due to presence of an exogenous mutagen and its reaction with the parent DNA, resulting in structural change that affects the base pairing capability of the altered nucleotide.
Mutations can be artificially induced with the help of two broad categories of mutagens-
a) Physical Mutagens: They are mainly radiations, although change in pH value, temperature can lead to induced mutation. The effect of a mutagen depends on its wavelength and penetrance. Lower is the wavelength, higher the penetrance.
b) Chemical Mutagens: Besides radiations, chemicals also help in inducing mutation. C. Auerbach was the first to detect mutations caused by chemicals as during World War II, chemicals used for inducing mutations were mustard gas, ethyl urethane, formalaldehyde and phenol.
Mutation at biochemical level.
Genes control various traits in an organism by controlling synthesis of proteins and expressing themselves through synthesis of enzymes demonstrated by G.W. Beadle and E.L.Tatum due to their discovery of biochemical mutation in Neurospora and proposed one gene one hypothesis. The hypothesis states that if in a biosynthetic pathway several steps are involved, each step will be controlled by a specific enzyme, which is synthesized under a control of specific gene.
Inborn born Errors of Metabolism in Man: Inborn Error of Metabolism are a group of disorders which are genetically transmitted (autosomal recessive or X linked recessive) and cause a single gene defect by blocking significantly the metabolic pathway. There are three important diseases associated with Inborn Errors of Metabolism namely Phenylketonuria, Alkaptonuria and Albinism. Phenylketonuria is due to accumulation of phenylpyruvic acid and causes mental retardation. Children suffering from Phenylketonuria are called Phenylpyruvic idiots and are thus unable to break Phenylpyruvic acid into hydroxyl phenylpyruvic into hydroxyl- phenylpyruvic acid. Alkaptonuria is caused due to accumulation of homogentisticacid, the urine of patients suffering with this disease turns black in contact of air.Albinism is due to the presence of melanin pigment and individuals suffering with this disease are incapable of converting dihydroxy phenylalanine into melanin.
BIOCHEMISTRY OF DEGRADATION OF PHENYLALANINE


Mutation at molecular level
Watson and Crick pointed out the structures of bases in DNA are not static. Hydrogen atoms move from one position in a purine or pyrimidine to another position, from an amino group to ring nitrogen. Such chemical fluctuations are called tautomeric shifts. Mutationsat the molecular level havepermanent alterations in sequences of nucleotides in the nucleic acid, which adds to the genetic material. These alterations in the genetic material can be of following types:
I. Deletion of bases
II. Insertion of bases
III. Inversion of a sequence
IV. Replacement of base pair
Deletion and insertion of base pairs lead to breakage and reunion of DNA segments.Base pairs addition and deletion are collectively known as frame-shift mutation because they alter the reading frame of all base pair triplets. In majority case, these results in a failure to synthesize a functional protein allowing the mutation to be identified by its phenotypic consequences.
FRAMESHIFT MUTATION IN A SINGLE BASE PAIR TO STRUCTURAL GENE

TABLE 1-DISEASES WITH THEIR TRIPLET REPEAT EXPANSION

Base pair replacement or substitution can be of two types namely Transition and Transversion. Transition comprises the substitution of one pyrimidine by the other or of one purine by the other. If a purine is replaced by a pyrimidine or replacement of pyrimidine by a purine is termed as transversion.Fourdifferent transition and eight different transversion are possible
Alternation in Base pairs of DNA sequences

Effect of chemical mutations Nucleotide sequences:
Chemical mutations effect nucleotide sequences in three ways
- By acting as a base analog, where the structural similarity to one of the bases in DNA enables it to act as mutagen.
- By reacting directly with the nitrogenous base, causing structural changes in the sequence.
- By reacting indirectly on DNA and causing the cells to synthesize chemicals having mutagenic effect.
Most common types of chemical mutation and their mode of action are described as below–
Base Analog-These are certain bases that are not present in DNA but bear a strong structural resemblance to normal nitrogenous bases and can be incorporated from the appropriate triphosphate precursor during DNA sequences.

Figure2:http://academic.pgcc.edu/~kroberts/Lecture/Chapter%207/0723_NucleotideAnalogs_L.jpg
{kind=link}
Figure 2 shows 5- bromouracil (5-Bu) having the same base pairing properties as thymine, Occasionally 5-Bu loses a hydrogen atom in position 1 and pairs with guanine instead of adenine. The mutagenic effect arises because the equilibrium between the two 5-bromouracil is shifted more towards enol formand leads to base pair substitution in daughter DNA molecules. The higher frequency of pairing mistakes is due to higher electronegativity of Br in Bu as compared to methyl group in thymine. The pyrimidine ring gets poorer in electrons and thus can lose H-atom at position 1 more easily than in thymine, thus giving rise to enol form
Inhibition of nucleic acid precursors– there are several agents which interfere with normal syntheisis of nucleic acid precursors as well as with synthesis of purines and pyrimidines. Azaserine and Urethane are alkylating agents is one of the strongest mutagen inhibiting purine and pyrimidine synthesis respectively chemicals such as ethyl methanalesulfonate add groups in DNA molecules called alkylating agents. The effect of alkylation depends on the position at which the nucleotide is modifiedand the type of alkyl group that is added. Nitrous acid (HNO3) is a major mutant that acts on replicating as well as non- replicating DNA and it results in oxidative deamination of the amino groups in adenine, guanine and cytosine.

Figure:https://learning.uonbi.ac.ke/courses/SZL311/scormPackages/path_2/qqqqqqqqqq.JPG
{kind=link}
Oxidative deamination of bases in DNA (a) adenine to hypoxanthine, causing A:T→G:C Transitions
(b) cytosine to Uracil causing C:G→A:T transitions and (c) Guanine to Xanthine, which is not mutagenic. Nitrous acid (HNO3) induces transitions in both the directions.
Inhibition of Acridine dyes: Acridine dyes such as proflavin, Acridine or orange, and a whole series of related compounds induces frame shift mutation. The positively charged acridines intercalate or stack themselves between DNA base pairs

Figure: Acridine Dyes intercalate DNA Sequence http://biosiva.50webs.org/dam.ht8.jpg
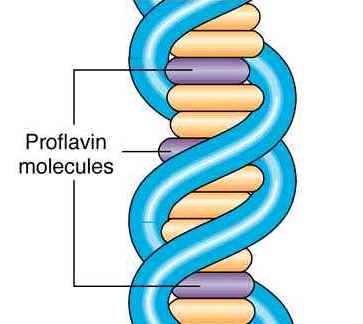{kind=link}
Deleterious Mutation
Many diseases are recognised in form dominant and recessive types, all of these are rare. Most of these diseases seem to be maintained in the population by a balance between selection and mutation.
Dominant Mutations as a cause of Disease
A particular type of dwarfism, called Achondroplasia is found at a very low frequency in nearly all human populations. The disease is not incompatible with life but dwarfs have a low chance of marrying and having children. However the trait is dominant and all affected individuals are heterozygous for the dominant dwarfing gene when one parent is chondrodystrophic and the other normal parent. The incidence at birth of achondroplasia is 1 in 10000. Natural selection keeps the frequency of this disease by eliminating homozygotes and by decreasing the chances of heterozygotes marrying and having children. In the absence of mutations, all affected individuals will have either an affected father or mother, because one of the two parents must have carried the dominant gene responsible for dwarfism. The example of Achondroplasia shows that mutation and natural selection maintains the low frequency of this disease. Natural selection acts not by death of a premature carrier, but through a relative inability to leave progeny, by increasing mortality rather than decreasing fertility. Production of new copies of the deleterious gene


The graph represents a hypothetical population which includes no achondroplastic dwarfs in the generation zero and the mutation rate is 1 per 20000 live births in each generation. Curve a represents the dwarfs reproducing at the same rate as done by normals there is a steady increase in the frequency of dwarfs as there is accumulation of new mutant genes in each generation. In second curve, natural selection counter balances the production of mutants and so there frequency in the population soon reaches a constant value which is higher than the mutation rate whereasthe incidence of the disease at birth stabilizes at a level that is somewhat higher than the mutation rate. Thus natural selection eliminates carriers of deleterious mutations, and prevents mutants beyond a certain incidence in the population.
Chromosomal Aberrations causing deleterious mutations
The phenotypes of many organisms are affected by changes in the number of chromosomes in their cells, these numerical changes are described as variation in the ploidy of organism. The existence of genetic variations involving aberration or defect in chromosomes.
Aberrations from Nondisjunction in Human Beings
Chromosomal Aberrations such as Down’s, Turner and Klinefelter’s syndrome are considered as deleterious dominants. Their fitness is close to zero and the estimates of mutation rate can be derived from their incidence of birth.Dominant genes are manifested in the form of a heterozygote and thus its effect can be escaped from selection and their frequency is low. High frequencies of recessive genes can be accumulated in the population with the help of mutation before natural selection acts to check their increase in frequency. For instance, the deleterious gene increases in the frequency by 1% as a result q=0.01 i.e. an incidence of this deleterious recessive gene is 1 among 100, thus the expected proportions of homozygous recessives is q2 = 0.0001 i.e. 1 in 10000. Due to adverse selection there is loss in the recessives genes because recessive homozygotes carry only recessive genes.
Genetic load
Concept and Definition
Genetic variability pertains to many advantages and disadvantages, many of the genes maintained by natural selection in a population are disadvantageous to it if their carriers are in homozygous state. The concept of genetic load was given by Haldane (1949) and Muller (1950). It hovers around an idea that the genotypes in a population have a mixture where one is better than the other. Genetic load is the difference between the fitness of an expected optimal genotype with the fitness of observed average genotype in a population. It accompanies with a loss of a portion of individual known as genetic death as there is a probability of an organism failing to reach the reproductive age. This fact is important to study by scientist as the population with too high genetic load will be in danger of going extinct as the organism are unable to survive and reproduce viable offspring. For instance if a gene is deleterious in homozygous state the frequency of homozygotes before and after selection will be as follows:

Thus the incurred genetic load by the individuals with aa genotype is sq2 , if there were 100 individuals in the population before selection, 100sq2 are eliminated because of genetic imperfection.
Haldane’s Dilemma
The value sq2 equals the mutation rate at equilibrium A→a, which is mutation cause by recessive deleterious gene, is equal to the mutation rate. On keeping the mutation rate constant a little difference can be seen in genetic load whether the Darwinian fitness s is small or large. High selection coefficient will lead to rapid elimination of genes (low q) whereas a low selection coefficient will let the gene to remain in the population (high q).
Causes of genetic load
Mutational load
Genetic loads are affected by mutation, which are negative as more mutations in a genome are deleterious than advantageous. In Crow terms Mutational load can be defined as the extent to which a population is impaired by recurrent mutation. Mutational load is larger in big populations with positive selection because there are more individuals that can be potential carrier of deleterious gene and can pass on the mutation over generations. In positive selection genome undergoes changes and also restores the mutation which is beneficial to the population. The mutational load has two components: one produced by deleterious mutation and the other produced by beneficial mutations. Both the components of mutational load arises because an ideal population consist of individual homozygous for both beneficial as well as deleterious gene.
Segregational Load
The segregational or the balancing load comes into play when heterozygote superiority arises in a population. For a gene with two alleles, segregational load can be computed,

By substituting the equilibrium frequency for q= s/s+t p=t/s+t is for the equation p2t+q2t is as follows-

Therefore if s and t both are equal to 1, the segregational load will be 0.05. this value is considered higher for all the mutation rate and demonstrates that in a randomly breeding population genetic load tends to increase by segregational load rather than mutational load.
Inbreeding
Inbreeding increases the rate of homozygosity, an inbreeding depression will be observed when the offspring’s receive two recessive and deleterious alleles resulting in a lower fitness. A declining population will experience high rate of mutation. An increase in the rate of mutation in alleles will increase the pairing frequency alleles. Due to a decline in population because of higher mutation rate, there is a chance of mating between the remaining organism to breed with their relatives and it will help in pushing out some of the harmful mutations
Sexual reproduction
Sexual reproduction has a direct effect on genetic load, it increases by pairing together of two different deleterious gene, referred to the concealed portion of genetic load. Sexual reproduction can also prevent the spread and maintenance of deleterious gene in a population due to elimination of less fit individuals. Genetic load will increase at a steady rate if mutations spreading to all offspring are slightly deleterious to the point where an individual can survive and reproduce.
Migration
Migrational load results when an organism non-adaptive to a particular environment comes in contact with individual adapted to that environment. The offspring produced as a result of such mating are not as fit as they would have been if both of their parent where adapted to same environment. The offspring will have a lower variability rate as well as a lower overall fitness and higher will be the genetic load if individuals are from remarkably different environment.
Case studies
Selectionsist pay attention on the association between the patterns of some enzyme poplymorphism and ecological conditions as human populations are usually affected by a number of heretiable disease which are merely caused by recessive genes. These effects are moreover rare in occurance. A strong correlation lies between the genes for sickle cell anaemia and malaria and also between the genes for thalassemia and malaria. A sharp difference lies between the manifestation of the disease, one is a qualitative whereas the other is the quantitative type of haemoglobinopathies. The geographical distribution of both genes offers protection against malaria. For instance Thalassaemia, a inherited autosomal recessive blood disorder where patients have in production of either alpha and beta chains of haemoglobin. The selective advantage is enjoyed fairly by over dominant heterozygote. Two forms of thalassemia, in the first one thalassemia major (homozygote for the defect) causes the red blood corpuscles to collapse and leading to severe anaemia which is fatal in childhood. The second form is thalassemia minor which is heterozygote for the trait. Immunity is attained and a selection advantage is achieved over normal. Incidence of the defective alleles will increase slightly if all heterozygotes can be persuaded to marry normal homozygotes then their children will also be normal.
Summary
- Mutations are rare, lethal, random, recessive to dominant and recurrent and can occur at any time and span of an individual’s lifetime.
- Mutation can occur in germinal cell, gamete or zygote and also in a cell (Somatic mutation) can occur at protein level( substitution of amino acid), at the DNA nucleotide level( substitution of one base for another, deletion of one base and insertion of new base) and at biochemical level.
- Point mutations occurring in non coding sequences are without consequences. Mutations occurring in coding sequences results in short and long term effects. The short term effect results in halting of the cell cycle causing production of deformed proteins whereas long term effect prohibits the cell from reproduction and thus lead to death of cell.
- Deamination a form of spontaneous lesions leads to damage in the naturally occurring DNA, the loss of the naturally occurring bases is in the presence of pH and temperature dependent reactions and results in the conversion of the affected bases into hyoxanthine, xanthine and thymine respectively.
- UV rays act as potent physical mutagen that increases the occurrence of mutation. H.J. Muller was the first to demonstrate that X rays can cause mutations, as an increased X-rays treatment increased the frequency of sex linked recessive lethal mutations in D. Melanogaster. The direct affects of chemical mutagens were the direct interaction of the radiation with DNA whereas indirect effect on the other side results from the interaction of DNA with reactive species formed by the radiation.
- The idea of genetic perfection with absence of genetic death is a myth and never achieved in a population as environmental conditions changes with time and also the advantages with different genotypes changes with time.
- A population with a relatively large genetic load is subjected to new environment where new deleterious gene survives.
- Thus genetic load can be measured in terms of departure from optimum genotype, population with too high genetic load will be in danger of getting extinct there is a reduction in the selective value for a population compared to a population with most favourable genotype.
| you can view video on Mutation And Genetic Load |
References
SUGGESTED READINGS
- Dobzhansky, T. (1977). Evolution. San Francisco: W.H.Freeman & Co.
- Hamilton, M. B. (2009). Population Genetics. Chischester: Wiley- blackwell.
- Pierce, B. A. (2013). Genetics: A Conceptual Approach. W.H.Freeman.
- Relethford, J. H. (2012). Human Population Genetics. new york: Willey Blackwell.
- Russell, p. J. (1997). genetics. Benjamin-Cummings Publishing Company; 5 Sub edition.
- Sforza, W. &. (1975). Genetics Evolution and Man. W H Freeman & Co .
- Strickberger, M. W. (2006). Genetics. South Asia: Pearson Prentice Hall.